The last two decades have seen important advances in our knowledge on protein folding and the stability of globular proteins, as a result of extensive studies using various biophysical approaches and theories. Calorimetry, particularly differential scanning calorimetry (DSC), has proven to be very useful for examining the thermodynamics of protein (un)folding and stability. A large number of DSC studies have revealed the general thermodynamic nature of thermal unfolding, i.e. cold and heat denaturation, by producing a series of thermodynamic parameters: ∆G, ∆H, ∆S, and ∆Cp, as well as the midpoint of unfolding temperature (TM). The signs of these parameters for thermal unfolding (i.e., positive values of ∆S and ∆Cp as well as negative values of ∆G and ∆H) are the same regardless of the type of protein; however, the value of each parameter differs from protein to protein. Stability curves for various proteins with increasing temperature, which are constructed using thermodynamic parameters and the Gibbs-Helmholtz equation, also explain how protein stability depends on solvent conditions, pressure, and mutations. Thus, DSC has also been successfully applied to the development of antibody drugs.
Advances in our understanding of the thermodynamics of protein folding have been achieved using DSC; however, knowledge on thermodynamic properties and molecular mechanisms leading to protein misfolding and aggregation is very limited due to the technical difficulties associated with the large size and heterogeneity of aggregates. Furthermore, the ability of calorimetric methods to investigate thermodynamics has not yet been applied to protein aggregation.
We herein concisely describe recent and unique applications of calorimetry (both DSC and ITC) to the examination of protein misfolding and aggregation. Observations of spontaneous and seeded amyloid formation, amyloid fibril associations, amorphous aggregation, and dissociation of aggregates using calorimetry are described. We also discuss calorimetry-based thermodynamic studies on protein aggregation and address structural features to characterize protein aggregates using the thermodynamic parameters of aggregation. These pioneering attempts are expected to promote calorimetry-based thermodynamic studies on protein misfolding and aggregation and may provide the basis for the development of treatments for and the prevention of aggregation-related diseases.
Unfolded proteins fold into thermodynamically-favorable native structures under the correct physiological conditions for gain of function (Figure 1) [1-4]. The burial of hydrophobic regions in a protein's interior, along with the exposure of hydrophilic residues to aqueous solutions stabilizes native structures and ensures solubility. Active native proteins perform their biological functions through highly regulated intermolecular interactions with their binding partners, under thermodynamic control.
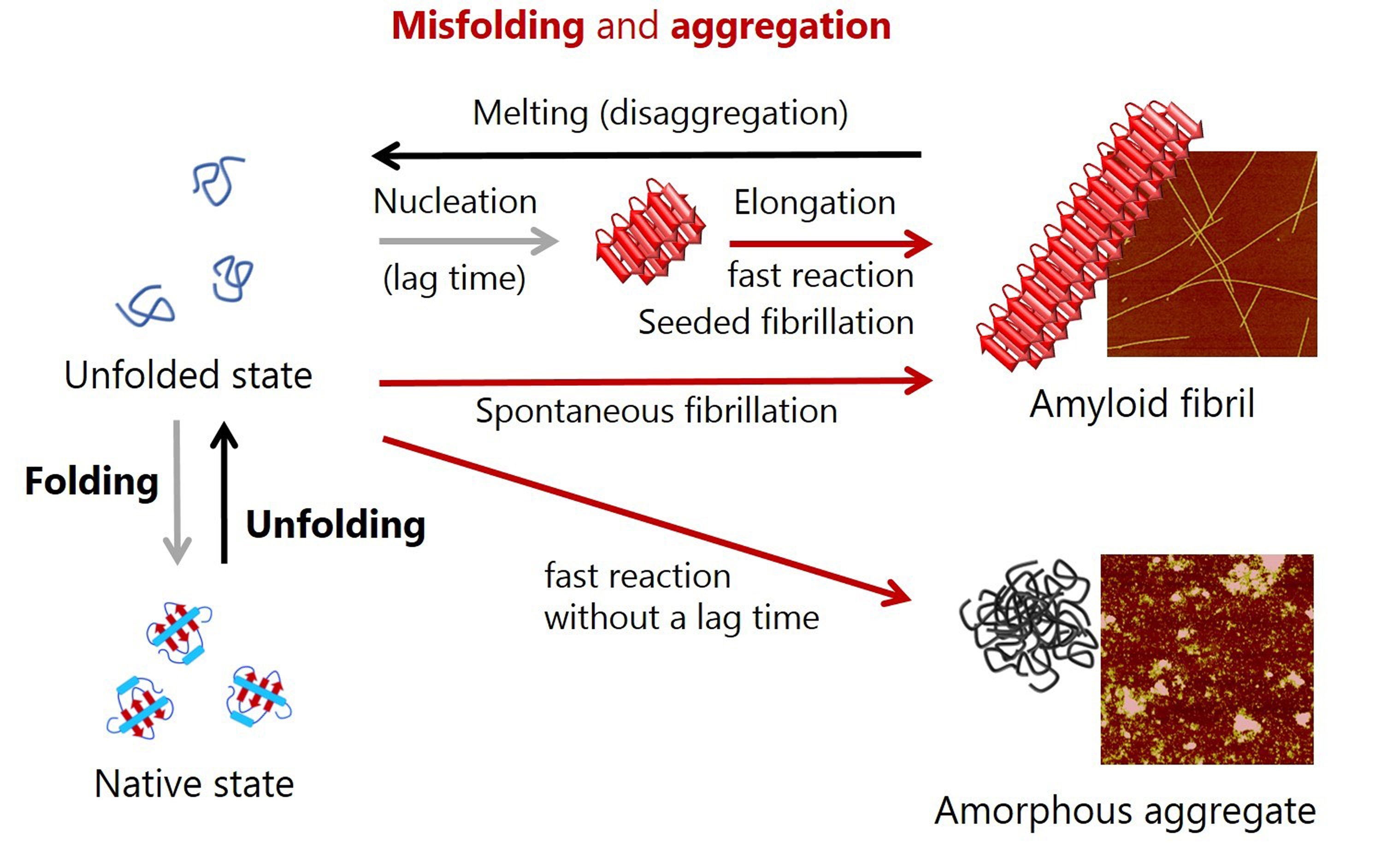
Figure 1: Schematic representation of conformational conversions by protein folding and protein misfolding-induced aggregation. Atomic force microscopy images of amyloid fibrils and amorphous aggregates are shown on the right.
Isothermal titration calorimetry (ITC) is a powerful approach for the thermodynamic investigation of interactions between diverse biomolecules and inorganic compounds [5-11] . ITC provides thermodynamic parameters for intermolecular interactions such as the affinity constant (i.e. dissociation constant ) ( K D ) as well as changes in Gibbs free energy (∆ G ), enthalpy (∆ H ), entropy (∆ S ), and heat capacity (∆ C p ), in addition to binding stoichiometry ( n ). Several key functions of the ITC instrument, such as the controlled titration of a sample in the ITC syringe (e.g., small molecules) into another sample in the ITC cell (e.g., macromolecules) and stirring, which ensures that the ITC cell solution is well suspended and mixed , further expand the utility of ITC to various binding systems.
Exogenous stress factors often deactivate native proteins and change thermodynamically stable states from native to locally or globally unfolded states with the exposure of aggregation-prone regions, which sequentially leads to the misfolding-coupled irreversible aggregation of proteins (loss of function). 'Quality control' in living biological systems plays a fundamental role in maintaining homeostasis by not only minimizing the deactivation and aggregation of proteins, but also maximizing protein turnover [12]. A failure to prevent aggregation and clear aggregates has been strongly implicated in the pathogenesis of a large number of diseases, including neurodegenerative diseases such as Alzheimer’s and Parkinson’s diseases, as well as amyloidosis [13,14]. The direct link between protein misfolding and aggregation and diseases has intensively promoted their study. These findings also suggest that protein folding, function, misfolding, and aggregation and disease are not separate events - they are closely related; therefore, a deeper understanding of each process and the interplay between them is critical.
Insoluble protein aggregates are classified into amyloid fibrils and amorphous aggregates [15-18]. Amyloid fibrils grow rapidly following a lag time which allows formation of productive nuclei by soluble precursor monomers, which is a nucleation-growth mechanism of amyloid formation (i.e., spontaneous amyloid fibrillation) (Figure 1) [13-18]. The addition of preformed amyloid fibrils to monomer solution omits the nucleation step, and, thus, only the elongation of amyloid fibrils (seeded amyloid fibrillation) is observed. Amyloid fibrils are ß structure-rich ordered protein aggregates that are morphologically straight, several mm long, and several nm in diameter [13,14]. In most cases, amorphous aggregation occurs very rapidly without an appreciable lag time (Figure 1) [15-18]. Amorphous aggregates do not have a particular appearance, but are often rounded. Other types of protein aggregates such as ß structure-containing oligomers and curved flexible protofibrils have been reported, but these share many of the properties of amyloid fibrils and amorphous aggregates.
The past two decades have seen important advances in our knowledge about protein folding and the stability of globular proteins, as a result of extensive studies using various biophysical approaches and theories [1-4,9,19-22]. Calorimetry, particularly differential scanning calorimetry (DSC), has proven very useful for examining the thermodynamics of protein (un)folding and stability. A large number of DSC studies have revealed the general thermodynamic nature of thermal unfolding, i.e. cold and heat denaturation, by producing a series of thermodynamic parameters: ∆G, ∆H, ∆S, and ∆Cp as well as the midpoint of unfolding temperature (TM) [3,9,19-22]. The signs of these parameters for thermal unfolding (i.e., positive values of ∆S and ∆Cp as well as negative values of ∆G and ∆H) are the same regardless of the type of protein; however, the value of each parameter differs from protein to protein. Stability curves for various proteins over temperature, which are constructed using thermodynamic parameters and the Gibbs-Helmholtz equation, also explain how protein stability depends on solvent conditions, pressure, and mutations. Thus, DSC has been also successfully applied to the development of antibody drugs.
Advances in our understanding of the thermodynamics of protein folding have been achieved using DSC; however, knowledge of the thermodynamic properties and molecular mechanisms leading to protein misfolding and aggregation is very limited, due to the technical difficulties associated with the large size and heterogeneity of aggregates. Furthermore, the ability of calorimetric methods to investigate thermodynamics has not yet been applied to protein aggregation.
We herein concisely describe the recent and unique applications of calorimetry (DSC and ITC) to the study of protein misfolding and aggregation. Observations of spontaneous and seeded amyloid formation, amyloid fibril associations, amorphous aggregation, and dissociation of aggregates using calorimetry have been described. We also discuss calorimetry-based thermodynamic studies of protein aggregation and address structural features to characterize protein aggregates using the thermodynamic parameters of aggregation. These pioneering attempts are expected to promote calorimetry-based thermodynamic studies on protein misfolding and aggregation and may provide the basis for the development of treatments for and the prevention of aggregation-related diseases.
DSC has been extensively used in thermodynamic examinations of the stability and thermal unfolding mechanisms of natively-folded proteins, based on characteristic transition peaks and well-established physicochemical relationships. Thermodynamic analysis of thermal unfolding accompanied by aggregation is often not possible when the occurrence of protein aggregates causes severe distortions in the Cp curve and irreversibility for unfolding.
However, modifications to and the deformation of DSC peaks and Cp traces may be used as indicators of the formation of aggregates (Figure 2), which are particularly important for verifying existing molecular species and the homogeneity of sample solutions, as well as elucidating the mechanisms underlying protein (un)folding. In 1998, Litvinovich and colleagues reported that consecutive DSC scanning of the recombinant module III-9 from murine fibronectin (rmIII-9) at neutral pH revealed the formation of multimers and aggregates of rmIII-9 with a broadened DSC peak and a deformed Cp curve with exothermic responses after the thermal transition (Figure 2A) [23].
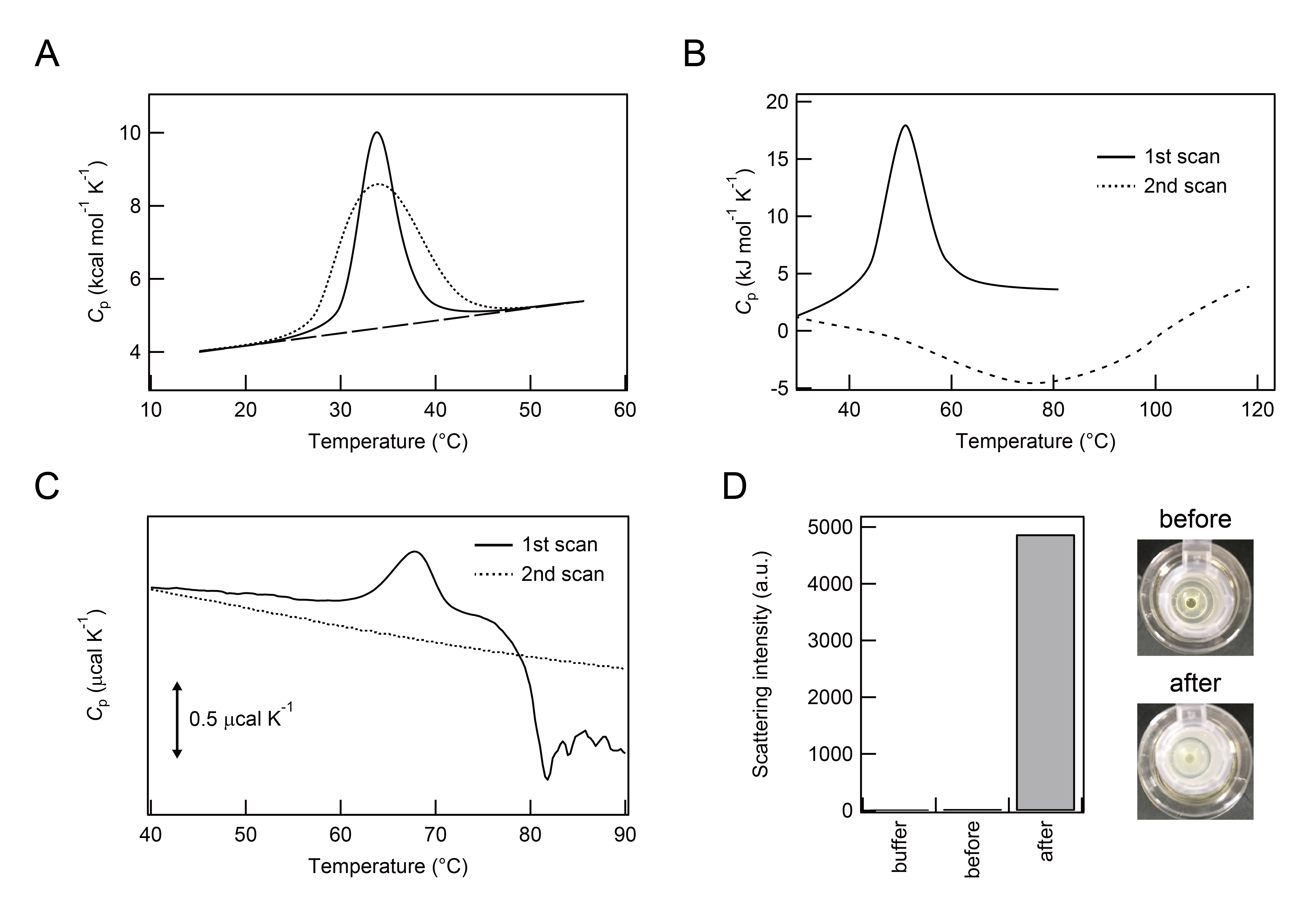
Figure 2: Thermal unfolding and subsequent aggregation of proteins. Panes A and B: Illustration of DSC thermograms of native folding and aggregation. Pane A: The continuous line represents a Cp curve of rmIII-9 observed and the dotted line indicates a best fit of the peak to a single two-state thermal transition (see Figure 5B of Litvinovich et al. [23] for more details). Pane B: First and second heating of ARQ are shown with continuous and dotted lines, respectively (see Figures 3C and 4D of Rezaei et al. for ARQ [25] for more details). Pane C: DSC thermograms of FNR. First and second heat scans are shown with continuous and dotted lines, respectively. Pane D: Top views of test tubes containing FNR sample solutions before (upper) and after (lower) DSC measurements on the right. Light scattering intensities of 25 mM sodium phosphate buffer (pH 7.4) alone, and FNR samples before and after DSC measurements on the left.
In 2002, Conejero-Lara and colleagues also interpreted similar aggregation-related thermal responses in DSC thermograms [24]. Deformed Cp curves for the thermal unfolding of streptokinase at mildly acidic pH or high protein concentrations were used to reveal the occurrence of aggregation and aggregation-induced irreversible thermal unfolding. They argued that kinetic controls (i.e. a scanning rate dependence) due to the slow exchange between thermally-unfolded soluble streptokinase and aggregates hampered the canonical thermodynamic DSC analysis. However, by fitting the DSC profiles to model equations, several thermodynamic parameters of streptokinase aggregation such as ∆H and ∆∆G from unfolded states to aggregation states were obtained.
In the same year, Rezaei et al. performed DSC studies on the relationship between disease susceptibility/resistance and the stabilities of several variants of the prion protein (PrP), which is closely related to neurodegenerative pathologies including prion diseases and transmissible spongiform encephalopathies [25]. Heat-induced unfolding of ARQ, one of the disease-susceptible variants, was irreversible due to the generation of fibrillar aggregates after repeated DSC scans (i.e. repeating the cycle of heating and subsequent cooling down) (Figure 2B). The second DSC up-scan showed exothermic peaks without an endothermic peak. Although the authors did not explain the origins of heat-generating processes in detail, the formation of fibrillar aggregates following the first DSC scan may be ascribed to these exothermic transition peaks (see below for more details). Thus, DSC thermograms with exothermic responses may be used to identify the presence of aggregates and/or aggregation.
In order to examine the origins of exothermic peaks, we performed DSC measurements of ferredoxin NADP+ reductase (FNR), which is a large (~35.5 kDa) multi-domain enzyme [11]. The first DSC scan of FNR also showed an exothermic response following an endothermic DSC peak for thermal unfolding, while the second scan did not display a characteristic peak in the Cp curve (Figure 2C). The sample solution after DSC measurements was cloudy and exhibited a high intensity of light scattering due to the formation of aggregates (Figure 2D), suggesting a correlation between an exothermic pattern of the Cp curve after thermal denaturation of proteins and protein aggregation/precipitation.
Clearer DSC profiles for amyloid formation were revealed by Dzwolak and Winter and their colleagues in 2003 and 2005 [26,27]. They unambiguously showed that amyloid formation by insulin, which causes injection-related amyloidosis, was accompanied by an exothermic transition after the endothermic thermal unfolding of native folds regardless of changes in experimental conditions (Figure 3) [26,27]. The Cp curves of insulin amyloid formation depended on the scan rate (i.e., kinetic controls) (Figure 3A), temperature (Figure 3B), and protein (Figure 3C) and alcohol concentrations (Figure 3D). The apparent ∆H for insulin amyloid formation at varying temperatures from 60 to 75 °C and ethanol concentrations from 0 to 30% were -16 – -57 kJ mol-1 (Figure 3B) and -70 – -140 kJ mol-1 (Figure 3D), respectively.
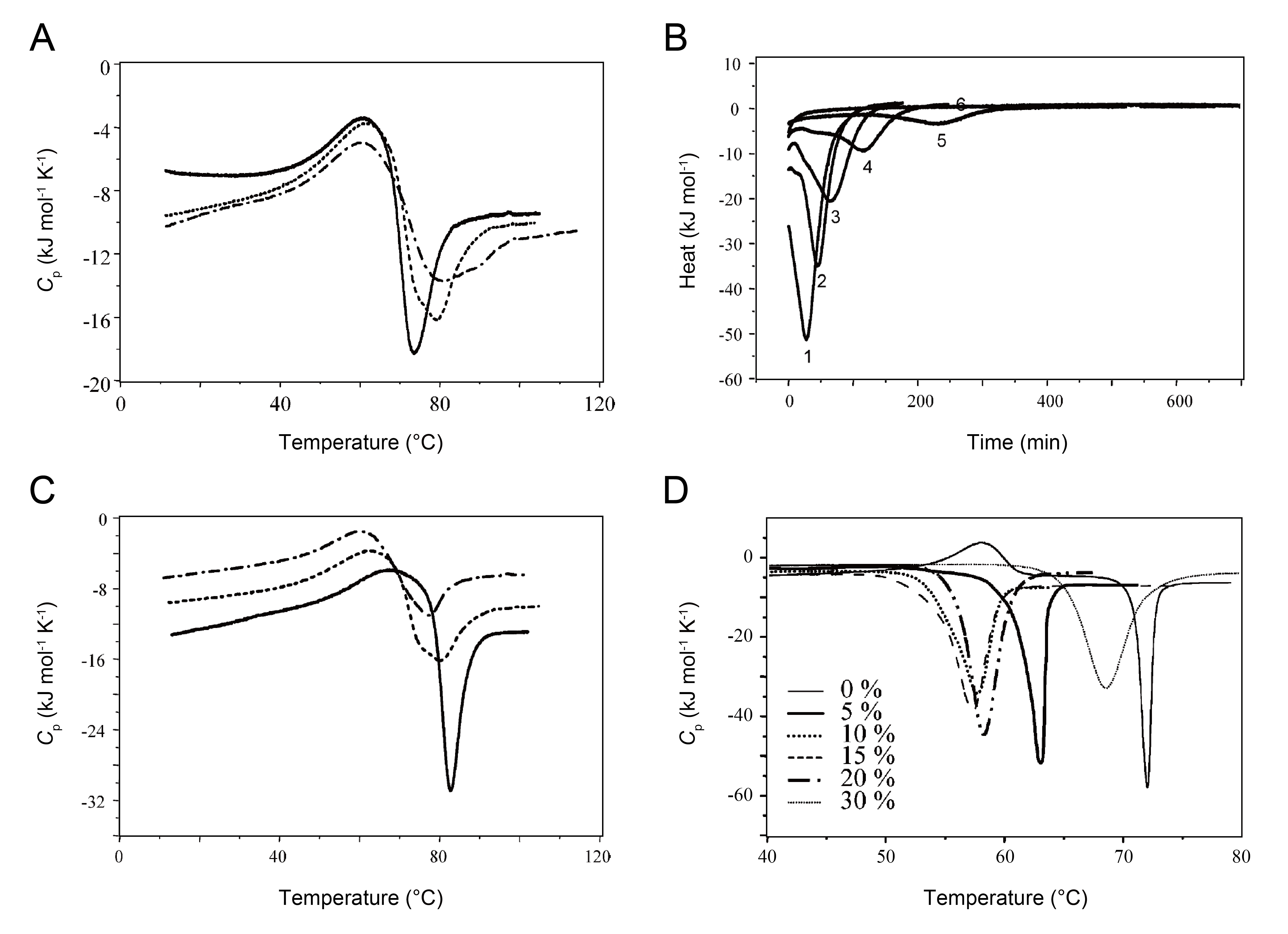
Figure 3: DSC scans of insulin aggregation at pH 1.9 under various experimental conditions. (A) DSC thermograms of insulin at distinct scan rates (10 (solid line), 20 (dotted line), and 10 °C h-1 (dashed dotted line)) [26]. (B) Heat obtained at 80 (curve 1), 75 (curve 2), 70 (curve 3), 65 (curve 4), 60 (curve 5), and 55 °C (curve 6) [26]. (C) DSC profiles obtained at 1 (solid line), 2 (dotted line), and 3% (dash dot line) with a scan rate of 20 °C h-1[26]. (D) DSC curves of 0.5 wt % insulin obtained at various ethanol concentrations, which are indicated in the figure [27].
Sasahara et al. conducted a series of DSC-based aggregation studies using several amyloidogenic proteins and peptides between 2005 and 2009 (Figure 4) [28-32]. They investigated two kinds of apparent exothermic heat: (1) the seed-dependent amyloid formation of β2-microglobulin (β2-m) responsible for dialysis-related amyloidosis (Figure 4A) [29, 32] and (2) amyloid fibrils of β2-m and amyloid β (Aβ) peptides/fragments, which cause Alzheimer’s disease (Figure 4B) [28].
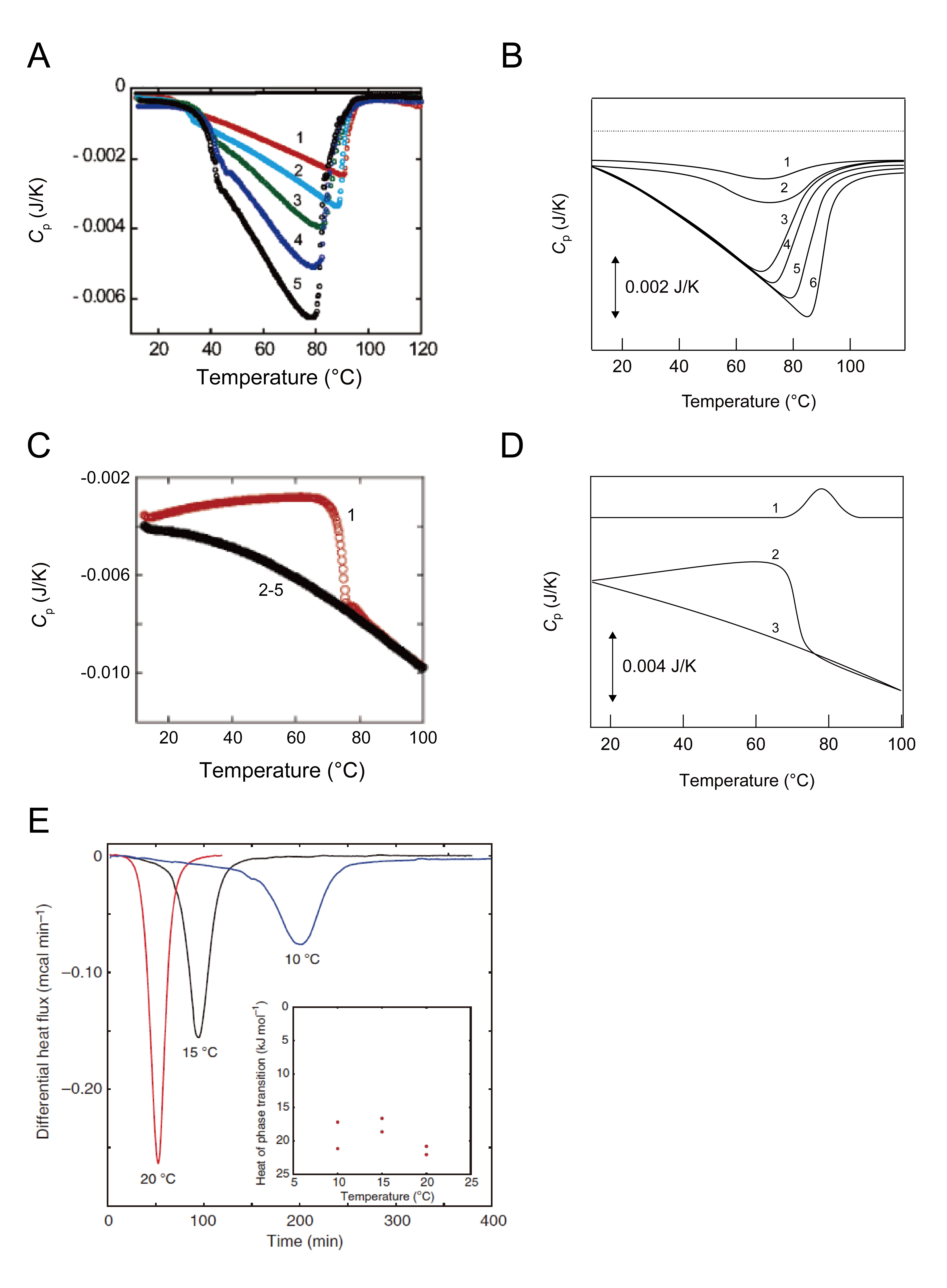
Figure 4: DSC scans for monitoring heat responses of amyloid fibrillation, amyloid fibrils, and conversion of aggregation states. Pane A: DSC thermograms of β2-m amyloid formation at various scan rates (20 °C h-1 (curve 1), 30 °C h-1 (curve 2), 50 °C h-1 (curve 3), 70 °C h-1 (curve 4), and 90 °C h-1 (curve 6)) with 0.2 mg mL-1 acidic-denatured β2-m and 0.5 mg mL-1 β2-m seed fibrils. The upper flat black curve indicates second heating after first heating to 120 °C [29]. Pane B: Schematic illustration of DSC traces of the heating of β2-m or Aβ(1-40) amyloid fibrils at varying protein concentrations. Increases in the concentration of β2-m or Aβ(1-40) are shown with numbering from 1 to 6 (see Figures 1A (β2-m) and 6A (Aβ(1-40)) of Sasahara et al. [28] for more details). Pane C: DSC thermograms of β2-m protofibrils formed at 0.5 M NaCl at a scan rate of 60 °C h-1[31]. Pane D: DSC thermograms of HEWL amorphous aggregates were depicted (see Figure 5A of Sasahara et al. [32] for more details). Curve 1 indicates the Cp curve of native HEWL. The number in curves 2 and 3 indicates the order of DSC heat scans. (E) Heat flow of the transition from spherical to tubular aggregates of diphenylalanine at the desired temperature [37].
The complex DSC thermograms of amyloid formation were interpreted to reflect multiple processes (Figure 4A) [28-32]. The initial sigmoidal exothermic effect due to the growth of amyloid fibrils (1st step) was followed by a gradual decrease in Cp curves due to the association among amyloid fibrils (2nd step), which is a process under kinetic controls based on scan rate-dependent increases in exothermic effects (Figures 4A and B). After the minimum Cp value, a marked increase in Cp (i.e., endothermic reaction) for the melting of amyloid fibrils was sequentially detected (3rd step).
This unique interpretation was supported by similar negative DSC peaks and kinetic controls in the DSC thermograms of the preformed amyloid fibrils of β2-m and the Aβ peptide (Figure 4B) as well as the heat denaturation of amyloid fibrils observed using far-UV CD spectroscopy [28]. The 2nd and 3rd steps in the DSC thermograms of preformed amyloid fibrils were very similar to those of seed-dependent amyloid formation. The 2nd step was reversible and scan-rate dependent, which, in turn, assigned the 1st step to the formation of amyloid fibrils. In addition, the onset temperature for the melting of preformed amyloid fibrils detected by far-UV CD spectroscopy was consistent with the temperature for the minimum Cp value, which is the starting point for the 3rd step.
The exothermic nature of amyloid formation in the 1st step and endothermic melting in the 3rd step are understandable. The origin of the thermal responses for the exothermic effects observed in the 2nd step may include specific and transient interactions between amyloid fibrils, dehydration [32], and the trapping of water, which decreases water motion [33].
Exothermic responses in DSC profiles due to the formation of α-crystalline amyloid fibrils were observed in consecutive scans [34]. Guzzi and colleagues also reported exothermic amyloid formation by β-lactoglobulin in the absence and presence of either Cu2+ or Zn2+[35,36].
Exothermic transitions have been reported between different types of aggregates [31,32,37]. Protofibrils of acid-denatured β2-m prepared by agitation at a high concentration of NaCl (0.5 M) were converted to mature amyloid fibrils as indicated by the dip in the Cp curve (i.e. an exothermic reaction) (Figure 4C) [31,32]. Amorphous aggregates of the native-like structures of β2-m at neutral pH and hen egg white lysozyme (HEWL) at pH 2-6 were transformed to mature amyloid fibrils, as shown by exothermic responses in DSC scans (Fig. 4D) [30,32].
Incubation in the DSC cell at a fixed temperature transformed the spherical assemblies of diphenylalanine, which may be regarded as amorphous aggregates, into highly-ordered tubular aggregates (Figure 4E) [37]. This phase transition also generated clear negative exothermic peaks with incubation times at several temperatures (i.e. isoscan mode in VP-DSC (MicroCalTM, Malvern Instruments, UK)).
In contrast to the exothermic reactions used to organize the structures of proteins by folding or misfolding-induced aggregation, unfolding or disaggregation may involve the uptake of heat. The disassembly of preformed amyloid fibrils showed positive endothermic DSC peaks [38-40]. The melting of amyloid fibrils formed from the N-terminal prion domain of Ure2p [38] and a variant of the α-spectrin SH3 domain, N47A [39,40] led to the absorption of heat (Figure 5A-C). Based on clear transition peaks, thermodynamic analyses were conducted. T
M
and ∆ H for the heat denaturation of Ure2p and N47A amyloids were ~75 °C and ~370 kJ mol -1 and ~75 °C – ~90 °C and ~100 kJ mol -1 – ~130 kJ mol -1 , respectively. Protofibrils formed from acid-denatured β2-m at 0.5 M NaCl were thermally depolymerized, as shown by a single endothermic peak (Figure 5D) [29,31] .
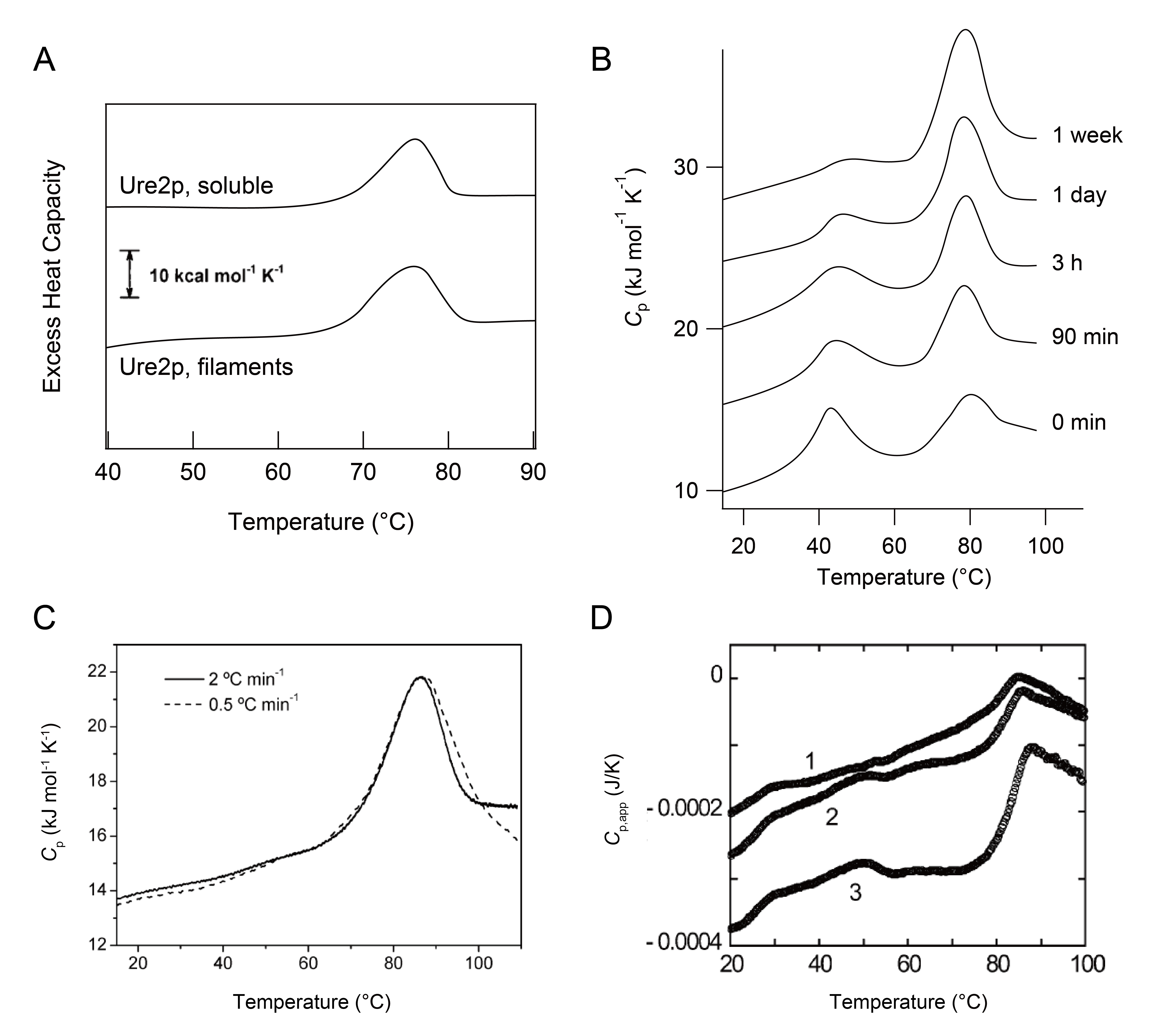
Figure 5: DSC scans showing the melting of protein aggregates. Pane A: DSC thermograms of soluble and aggregated Ure2p in 20 mM sodium phosphate buffer (pH 7.6) containing 100 mM NaCl. Samples were heated at 120 °C h-1[38]. Pane B: DSC curves of the N47A variant of the SH3 domain at 8.3 mg mL-1 in 100 mM glycine buffer (pH 3.2) containing 100 mM NaCl. Pre-incubation times are shown. Transition peaks at higher temperatures resulted from the melting of N47A amyloid fibrils [39]. Pane C: Cp curves of the amyloid fibrils of N47A at 5.5 mg mL-1 prepared in 100 mM glycine buffer for 1 month with distinct scan rates [40]. Pane D: Melting of β2-m protofibrils formed at 0.5 M NaCl scanned at 60 °C h-1[29]. β2-m concentrations were 0.2 mg mL-1 (curve 1), 0.3 mg mL-1 (curve 2), and 0.5 mg mL-1 (curve 3). It should be noted that Figure 5A and B were reproduced based on original papers [38,39].
Several limitations associated with the application of DSC to the study of protein aggregation should be considered. Difficulties are often associated with accurately and precisely estimating the thermodynamic parameters of protein aggregation using DSC because there are constant fluctuations in temperature during aggregation, and the stability of aggregates and pathway of aggregation are dependent on temperature. DSC thermograms can become unreliable due to large noise and low reproducibility stemming from the precipitation of large protein aggregates and sticky aggregates, which may adhere to the surface of a DSC cell. Thus, the interpretation of kinetic thermal responses requires more careful analysis and more convincing evidence.
The exact mechanism underlying the unusual thermal response of protein aggregation may be elucidated through more systematic case studies using DSC. However, the findings described above provide us with a quick qualitative judgement for the formation and clearance of amyloid fibrils simply by observing negative and positive DSC peaks, respectively, beyond quantitative evaluations.
ITC, instead of DSC, may be used to investigate protein aggregation by use of its stirring function and controlled titration at a constant temperature. ITC may be expected to produce more reliable and reproducible data on protein aggregation than DSC because continuous stirring disperses aggregate solutions preventing precipitation, and thereby efficiently transfers reaction heat to thermal sensors. We showed that ITC represents a powerful approach for investigating protein aggregation and allows for the thermodynamic characterization of protein aggregation.
Limited information is available on ITC-based examinations of protein aggregation [41-43]. The first trial was conducted by Kardos and colleagues in 2004, who successfully detected exothermic heat produced during the elongation of β2-m amyloid fibrils using VP-ITC (MicroCalTM, Malvern Instruments, UK) and thermodynamically characterized protein aggregation (Figure 6A and B) [41]. Preformed β2-m amyloid fibrils (i.e. seed fibrils) in the ITC syringe were titrated into acid-denatured β2-m monomers in the ITC cell for the single monitoring of the fibril growth reaction (Figure 6A). Alternatively, acid-denatured β2-m monomers in the ITC syringe were titrated into β2-m seed fibrils in the ITC cell for repeated observations of fibril growth reactions in a single ITC experiment, because individual injections induce fibril elongation. The ∆H value obtained at 37 °C was 124 kJ mol-1 and varied in a temperature-dependent manner. The ∆Cp value for β2-m fibril growth was deduced from the temperature dependence of ∆H (Figure 6B). The ∆G and ∆S of amyloid fibrils were also estimated to be -43.9 kJ mol-1 and 80 kJ mol-1 at 37 °C, respectively, based on residual monomers under the two-state equilibrium between monomers and fibrils and the relationship, ∆G = ∆H - T∆S = -RTlnK, where R and K are the gas constant and equilibrium constant, respectively. Negative ∆H and positive ∆S were the driving forces for amyloid formation at 37 °C.
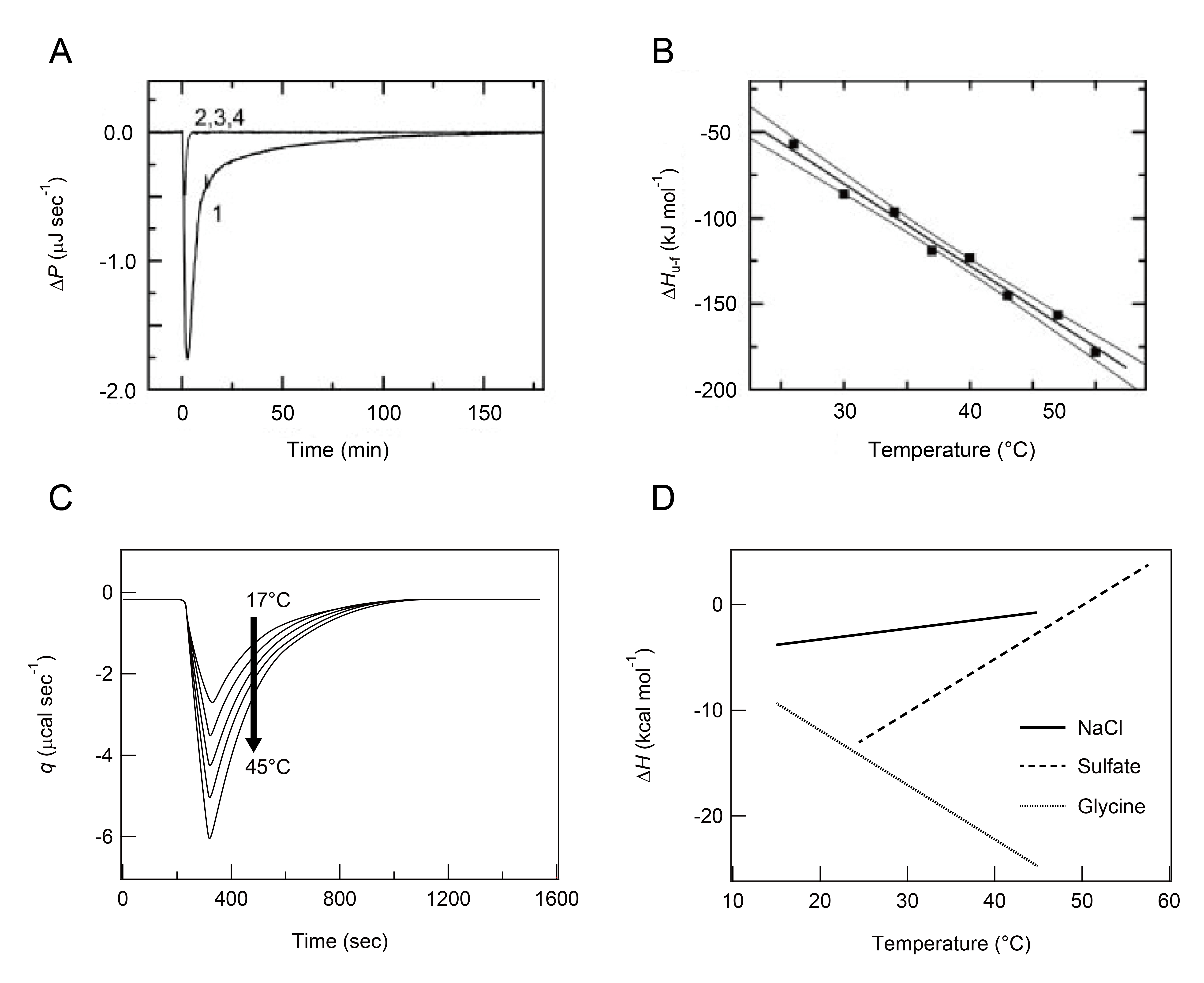
Figure 6: Observation of heat and temperature-dependent ∆H for the growth of amyloid fibrils using ITC. Pane A: ITC thermograms recorded by the injection of seeds at a final concentration of 15 μg mL-1 in the ITC syringe to a solution of β2m monomers at 0.1 mg mL-1 in the ITC cell at 37 °C (line 1). Overlapping thin lines indicate a series of reference measurements as buffer to buffer (line 2), buffer to monomer (line 3), and seed to buffer solution (line 4) [41]. Pane B: Each ∆H (black sphere) of the β2m fibril extension was plotted as a function of temperature. The black line indicates the best fit curve with a straight line [41]. Pane C: A schematic representation of ITC thermograms of the growth of glucagon amyloid fibrils in sulfate buffer containing 10 mM HCl and 1 mM Na2SO4, obtained by increasing temperatures from 17 °C to 45 °C (see Figure 1A of Jeppesen et al. [42] for more details). Pane D: Temperature dependence of ΔH measured in the three distinct buffers (5 mM HCl solution containing 150 mM NaCl (solid line), 50 mM glycine buffer (pH 2.5) containing 1 mM sulfate (broken line), and 50 mM glycine buffer (pH 2.5) (dotted line)) are schematically shown (see Figure 1B of Jeppesen et al. [42] for more details).
Jeppesen and colleagues also observed the heat of the elongation reaction of fibrils formed by glucagon, a peptide hormone, using the same ITC method (Figure 6C) [42]. They obtained ∆H and ∆Cp values for glucagon fibril growth and revealed the polymorphic nature of glucagon fibrillation in terms of the different thermodynamic responses (i.e. the sign and magnitude) of ∆H and ∆Cp depending on solvent conditions (i.e. types of salts), which showed another useful application for ITC besides thermodynamic investigations (Figure 6D) [42].
Ikenoue and Lee with their colleagues very recently succeeded in the real-time monitoring of the heat of spontaneous amyloid formation and amorphous aggregation of β2-m through their unique use of VP-ITC (MicroCalTM, Malvern Instruments, UK) (Figure 7) [44]. They reported that the calorimetry-based thermodynamic characterization of protein aggregation was possible with precision and accuracy, and characterized the distinct conformational states of natively folded and misfolding-induced aggregated states of β2-m.
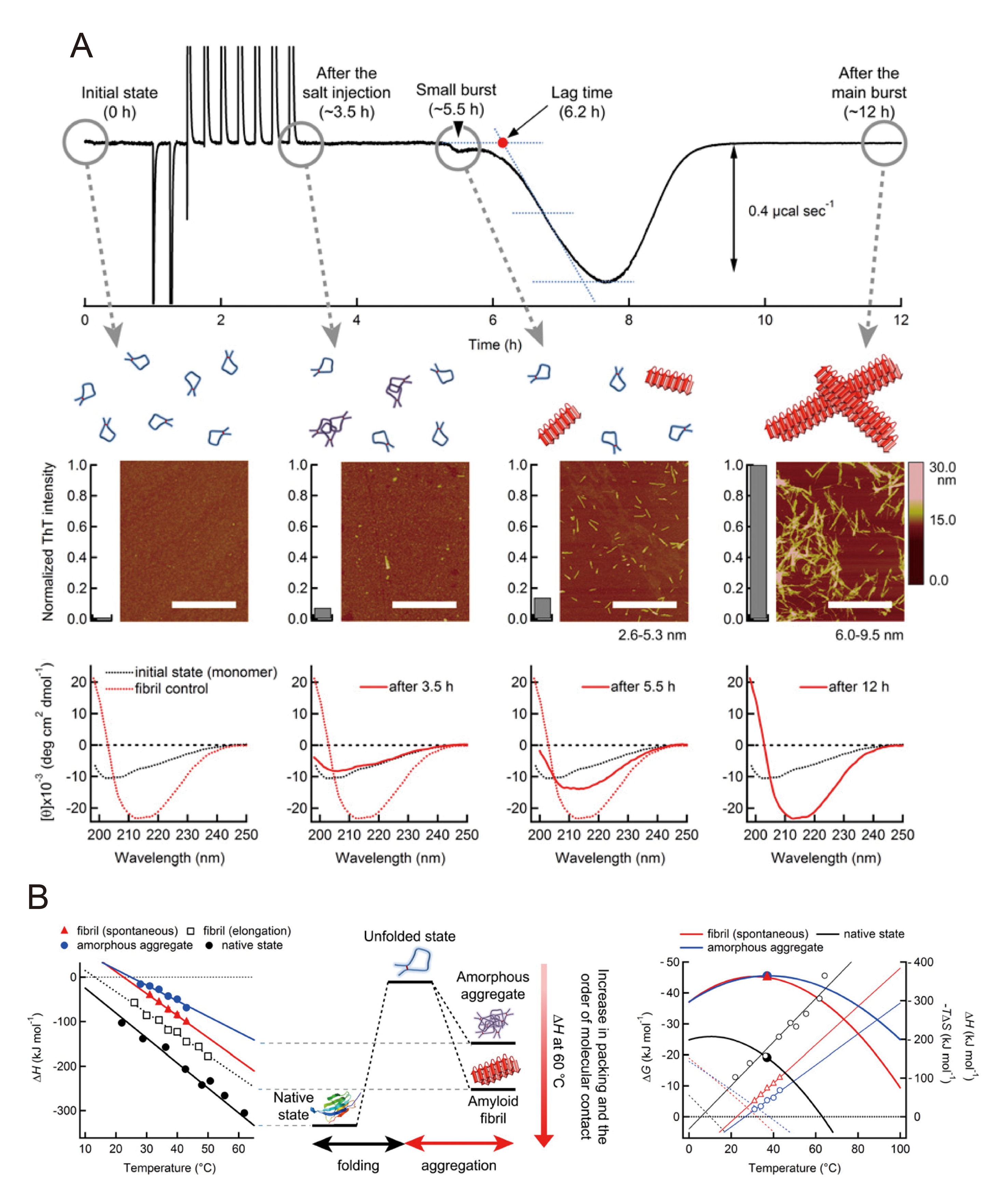
Figure 7: Monitoring the heat of the spontaneous amyloid formation of β2m using ITC and other approaches[44]. Pane A: The ITC trace of β2-m (1.1 mg mL-1) at pH 2.5 and 37 °C (upper panel). Conformational states of β2-m during an incubation in the ITC cell examined using AFM images and thioflavin T (ThT) fluorescence intensities (middle panel) and far-UV CD spectra (lower panel) at the four time points (0 h, ~3.5 h, ~5.5 h, and ~12 h). Illustrations of the molecular species of β2m based on AFM, ThT fluorescence, and CD are shown above AFM images: monomers (blue curves), prefibrillar aggregates (magenta curves), and amyloid fibrils (red rectangles). The CD spectra at each time point are shown by red solid curves. As a comparison, the spectra of monomers and mature fibrils (red dotted curves) are shown with black and red dotted curves, respectively. Pane B: Temperature-dependent ∆H for amorphous aggregation (blue circles), spontaneous (red triangles) and seed-dependent fibrillation (black squares), and folding (black circles) (left) are shown. The difference in ∆H at 60 °C among the distinct conformational states is shown with animation (right).
NaCl in the ITC syringe was gradually titrated to acid-denatured β2-m monomers in the ITC cell to reach a final concentration of 100 mM. Gradual increases in the NaCl concentration in the ITC cell decreased the solubility of β2-m monomers and changed the metastability of supersaturation, which produced a lag time. Large exothermic heat was followed by a lag time after salt titration (Figure 7A, upper panel). Exothermic fibrillation heat could be separated from the large endothermic peaks of the dilution heat of the salt injection due to the lag time. This large exothermic heat was attributable to the formation of amyloid fibrils, as supported by data from other approaches including a thioflavin T assay, far-UV CD spectroscopy, and atomic force microcopy (Figure 7A, middle and lower panels).
Continuous stirring as an effective agitation released β2-m monomers from the kinetically-trapped supersaturated states of β2-m by the metastability of supersaturation, thereby making effective nuclei for amyloid formation. ΔH at 37 °C was approximately -80 kJ mol-1. The magnitude of ΔH decreased with reductions in temperature, as observed for β2-m fibril growth, and ΔCp was also estimated from the slope of the ΔH plot against temperature (Figure 7B, left).
Reverse titration has been performed to evaluate the thermodynamics of β2-m amorphous aggregation at a high concentration of NaCl. The monomeric β2-m solution in the ITC syringe was injected into the ITC cell containing 1.0 M NaCl. Reaction heat was observed without an apparent lag time due to the low metastability of supersaturation, and amorphous aggregation was confirmed using several methods after reactions. ΔH at 37 °C was approximately -40 kJ mol-1 and ΔCp was also obtained from the linear temperature dependence of ΔH (Figure 7B, left). ΔG and ΔS were obtained in the same manner as seed-dependent amyloid fibrillation.
Based on the thermodynamic parameters obtained by ITC, we were also able to characterize the macroscopic conformational states of β2m in distinct conformations, i.e. natively folded states, amyloid fibrillar states, and amorphous aggregated states (Figure 7B, left), which were not amenable to other techniques. The DCp of the transitions of unfolded β2-m to three conformational states were all similar (~ -3.5 kJ mol-1 K-1 - ~ -5.5 kJ mol-1 K-1), which suggested a similar buried accessible surface area between β2-m folding and aggregation. Thus, ΔH for β2-m folding and aggregation was ascribed to differences in internal interactions.
The order of the magnitude of ∆H at 60 °C was: native folds (~ -300 kJ mol-1) > amyloid fibrils (~ -200 kJ mol-1) > amorphous aggregates (~ -120 kJ mol-1) (Figure 7B, right), suggesting that the internal packing of native β2-m obtained by optimized side-chain interactions was higher than that of the misfolded β2-m aggregates mainly formed by main-chain interactions. Highly ordered amyloid fibrils with internal hydrogen bond networks are more tightly packed states than amorphous aggregates, which do not have specific internal interactions.
Protein misfolding and aggregation are some of the central issues in various research fields. Powerful calorimetric methods have been applied to reveal the underlying mechanisms and thermodynamic properties of protein misfolding and aggregation. By maximizing the merits of DSC and ITC, more information on the thermodynamic behavior of abnormal aggregation is expected to be obtained. Although the thermodynamics of protein misfolding aggregation have not yet been completely elucidated, more case studies will contribute to establishing the thermodynamics of disease-related protein aggregates in parallel with the thermodynamics of protein folding.
In addition to detailed qualitative and quantitative evaluations on the thermodynamics of protein aggregation, heat itself is a simple, quick indicator for judging the formation and types of aggregates by observation of the sign and magnitude of the reaction heat on thermograms. These properties of the reaction heat of aggregation (sign and magnitude as well as the shape and pattern of the thermogram) will also be applicable to kinetics and the development of inhibitors of protein aggregation.
We thank Professor Yuji Goto (Osaka University, Japan) and Associate Professor József Kardos (Eötvös Loránd University, Hungary) for their comments on calorimetric studies. We thank Dr. Natalia Markova (Malvern Instruments, UK) for critical reading of this manuscript. We also thank Ms. Masako Hirose and Ms. Takuma Waki (Malvern Instruments, Japan) for their kind support of our calorimetric studies.
1 Fersht, A. R. and Daggett, V. (2002) Protein folding and unfolding at atomic resolution. Cell 108, 573-582
2 Daggett, V. and Fersht, A. (2003) The present view of the mechanism of protein folding. Nat Rev Mol Cell Biol 4, 497-502
3 Baldwin, R. L. (2007) Energetics of protein folding. J Mol Biol 371, 283-301
4 Nickson, A. A. and Clarke, J. (2010) What lessons can be learned from studying the folding of homologous proteins? Methods 52, 38-50
5 Jelesarov, I. and Bosshard, H. R. (1999) Isothermal titration calorimetry and differential scanning calorimetry as complementary tools to investigate the energetics of biomolecular recognition. J Mol Recognit 12, 3-18
6 Leavitt, S. and Freire, E. (2001) Direct measurement of protein binding energetics by isothermal titration calorimetry. Curr Opin Struct Biol 11, 560-566
7 Markova, N. and Hallen, D. (2004) The development of a continuous isothermal titration calorimetric method for equilibrium studies. Anal Biochem 331, 77-88
8 Velazquez Campoy, A. and Freire, E. (2005) ITC in the post-genomic era...? Priceless. Biophys Chem 115, 115-124
9 Privalov, P. L. (2009) Microcalorimetry of proteins and their complexes. Methods Mol Biol 490, 1-39
10 Ladbury, J. E. (2010) Calorimetry as a tool for understanding biomolecular interactions and an aid to drug design. Biochem Soc Trans 38, 888-893
11 Lee, Y. H., Ikegami, T., Standley, D. M., Sakurai, K., Hase, T. and Goto, Y. (2011) Binding energetics of ferredoxin-NADP+ reductase with ferredoxin and its relation to function. Chembiochem 12, 2062-2070
12 Powers, E. T., Morimoto, R. I., Dillin, A., Kelly, J. W. and Balch, W. E. (2009) Biological and chemical approaches to diseases of proteostasis deficiency. Annu Rev Biochem 78, 959-991
13 Lee, Y. H. and Goto, Y. (2012) Kinetic intermediates of amyloid fibrillation studied by hydrogen exchange methods with nuclear magnetic resonance. Biochim Biophys Acta 1824, 1307-1323
14 Knowles, T. P., Vendruscolo, M. and Dobson, C. M. (2014) The amyloid state and its association with protein misfolding diseases. Nat Rev Mol Cell Biol 15, 384-396
15 Yoshimura, Y., Lin, Y., Yagi, H., Lee, Y. H., Kitayama, H., Sakurai, K., So, M., Ogi, H., Naiki, H. and Goto, Y. (2012) Distinguishing crystal-like amyloid fibrils and glass-like amorphous aggregates from their kinetics of formation. Proc Natl Acad Sci U S A 109, 14446-14451
16 Lin, Y., Lee, Y. H., Yoshimura, Y., Yagi, H. and Goto, Y. (2014) Solubility and supersaturation-dependent protein misfolding revealed by ultrasonication. Langmuir 30, 1845-1854
17 Muta, H., Lee, Y. H., Kardos, J., Lin, Y., Yagi, H. and Goto, Y. (2014) Supersaturation-limited amyloid fibrillation of insulin revealed by ultrasonication. J Biol Chem 289, 18228-18238
18 Lin, Y., Kardos, J., Imai, M., Ikenoue, T., Kinoshita, M., Sugiki, T., Ishimori, K., Goto, Y. and Lee, Y. H. (2016) Amorphous Aggregation of Cytochrome c with Inherently Low Amyloidogenicity Is Characterized by the Metastability of Supersaturation and the Phase Diagram. Langmuir 32, 2010-2022
19 Privalov, P. L. and Dragan, A. I. (2007) Microcalorimetry of biological macromolecules. Biophys Chem 126, 16-24
20 Privalov, P. L. (2007) Reflections on the origins of microcalorimetry of biopolymers. Biophys Chem 126, 13-15
21 Iida, T., Nishimura, S., Mochizuki, M., Uchiyama, S., Ohkubo, T., Urade, Y., Tanaka, A. and Inui, T. (2008) Thermal unfolding mechanism of lipocalin-type prostaglandin D synthase. FEBS J 275, 233-241
22 Schon, A. and Freire, E. (2016) Three easy pieces. Biochim Biophys Acta 1860, 975-980
23 Litvinovich, S. V., Brew, S. A., Aota, S., Akiyama, S. K., Haudenschild, C. and Ingham, K. C. (1998) Formation of amyloid-like fibrils by self-association of a partially unfolded fibronectin type III module. J Mol Biol 280, 245-258
24 Azuaga, A. I., Dobson, C. M., Mateo, P. L. and Conejero-Lara, F. (2002) Unfolding and aggregation during the thermal denaturation of streptokinase. Eur J Biochem 269, 4121-4133
25 Rezaei, H., Choiset, Y., Eghiaian, F., Treguer, E., Mentre, P., Debey, P., Grosclaude, J. and Haertle, T. (2002) Amyloidogenic unfolding intermediates differentiate sheep prion protein variants. J Mol Biol 322, 799-814
26 Dzwolak, W., Ravindra, R., Lendermann, J. and Winter, R. (2003) Aggregation of bovine insulin probed by DSC/PPC calorimetry and FTIR spectroscopy. Biochemistry 42, 11347-11355
27 Dzwolak, W., Grudzielanek, S., Smirnovas, V., Ravindra, R., Nicolini, C., Jansen, R., Loksztejn, A., Porowski, S. and Winter, R. (2005) Ethanol-perturbed amyloidogenic self-assembly of insulin: looking for origins of amyloid strains. Biochemistry 44, 8948-8958
28 Sasahara, K., Naiki, H. and Goto, Y. (2005) Kinetically controlled thermal response of beta2-microglobulin amyloid fibrils. J Mol Biol 352, 700-711
29 Sasahara, K., Naiki, H. and Goto, Y. (2006) Exothermic effects observed upon heating of beta2-microglobulin monomers in the presence of amyloid seeds. Biochemistry 45, 8760-8769
30 Sasahara, K., Yagi, H., Naiki, H. and Goto, Y. (2007) Heat-induced conversion of beta(2)-microglobulin and hen egg-white lysozyme into amyloid fibrils. J Mol Biol 372, 981-991
31 Sasahara, K., Yagi, H., Naiki, H. and Goto, Y. (2007) Heat-triggered conversion of protofibrils into mature amyloid fibrils of beta2-microglobulin. Biochemistry 46, 3286-3293
32 Sasahara, K., Yagi, H., Naiki, H. and Goto, Y. (2009) Thermal response with exothermic effects of beta2-microglobulin amyloid fibrils and fibrillation. J Mol Biol 389, 584-594
33 Bergqvist, S., Williams, M. A., O'Brien, R. and Ladbury, J. E. (2004) Heat capacity effects of water molecules and ions at a protein-DNA interface. J Mol Biol 336, 829-842
34 Attanasio, F., Cataldo, S., Fisichella, S., Nicoletti, S., Nicoletti, V. G., Pignataro, B., Savarino, A. and Rizzarelli, E. (2009) Protective effects of L- and D-carnosine on alpha-crystallin amyloid fibril formation: implications for cataract disease. Biochemistry 48, 6522-6531
35 Stirpe, A., Rizzuti, B., Pantusa, M., Bartucci, R., Sportelli, L. and Guzzi, R. (2008) Thermally induced denaturation and aggregation of BLG-A: effect of the Cu(2+) and Zn (2+) metal ions. Eur Biophys J 37, 1351-1360
36 Guzzi, R., Rizzuti, B., Labate, C., Zappone, B. and De Santo, M. P. (2015) Ferric Ions Inhibit the Amyloid Fibrillation of beta-Lactoglobulin at High Temperature. Biomacromolecules 16, 1794-1801
37 Levin, A., Mason, T. O., Adler-Abramovich, L., Buell, A. K., Meisl, G., Galvagnion, C., Bram, Y., Stratford, S. A., Dobson, C. M., Knowles, T. P., et al. (2014) Ostwald's rule of stages governs structural transitions and morphology of dipeptide supramolecular polymers. Nat Commun 5, 5219
38 Baxa, U., Ross, P. D., Wickner, R. B. and Steven, A. C. (2004) The N-terminal prion domain of Ure2p converts from an unfolded to a thermally resistant conformation upon filament formation. J Mol Biol 339, 259-264
39 Morel, B., Casares, S. and Conejero-Lara, F. (2006) A single mutation induces amyloid aggregation in the alpha-spectrin SH3 domain: analysis of the early stages of fibril formation. J Mol Biol 356, 453-468
40 Morel, B., Varela, L. and Conejero-Lara, F. (2010) The thermodynamic stability of amyloid fibrils studied by differential scanning calorimetry. J Phys Chem B 114, 4010-4019
41 Kardos, J., Yamamoto, K., Hasegawa, K., Naiki, H. and Goto, Y. (2004) Direct measurement of the thermodynamic parameters of amyloid formation by isothermal titration calorimetry. J Biol Chem 279, 55308-55314
42 Jeppesen, M. D., Hein, K., Nissen, P., Westh, P. and Otzen, D. E. (2010) A thermodynamic analysis of fibrillar polymorphism. Biophys Chem 149, 40-46
43 Ikenoue, T., Lee, Y. H., Kardos, J., Saiki, M., Yagi, H., Kawata, Y. and Goto, Y. (2014) Cold denaturation of alpha-synuclein amyloid fibrils. Angew Chem Int Ed Engl 53, 7799-7804
44 Ikenoue, T., Lee, Y. H., Kardos, J., Yagi, H., Ikegami, T., Naiki, H. and Goto, Y. (2014) Heat of supersaturation-limited amyloid burst directly monitored by isothermal titration calorimetry. Proc Natl Acad Sci U S A 111, 6654-6659